过氧化氢(H2O2)触发的信号调控是通过协调全基因组转录重编程来调节植物的生长和防御。但是,控制H2O2依赖性基因表达的具体机制仍知之甚少。在这里,研究人员确定拟南芥介体复杂的亚基MED8作为H2O2响应的调节剂。在组成型氧化应激遗传背景(过氧化氢酶缺陷型,cat2)中引入med8突变与水杨酸途径的活性增强和细胞死亡加速相关。有趣的是,med8幼苗对除草剂甲基紫精产生的氧化胁迫具有更高的耐受性,并表现出防御信号的转录超活化,特别是水杨酸和茉莉酸相关的途径。med8触发的对甲基紫精的耐受性是通过在水杨酸和茉莉酸途径中引入二级突变来控制的。此外,对介体相互作用基因组的分析揭示了与参与mRNA加工和microRNA生物发生的成分的相互作用,因此将介体的作用扩展到了转录之外。值得注意的是,MED8与转录调节子NEGATIVE ON TATA-LESS(NOT2)相互作用,以控制H2O2诱导型基因的表达和应激反应。本文的工作建立了MED8作为调节氧化应激反应的成分,并证明它作为H2O2驱动的防御基因表达激活的负调节剂。
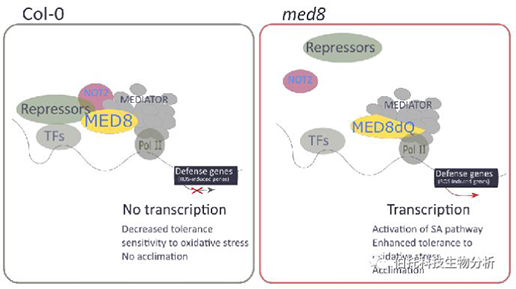
MED8在氧化应激信号传导中发挥作用的假设机理
在野生型背景下,全长MED8与NOT2以及其他未知的“阻遏物”相关联,从而抑制防御基因的表达并因此降低了对氧化应激的耐受性。在med8背景下,缺少MED8 C端区域可能会导致其与阻遏物模块的分离,防御基因的表达以及植物激素相关途径(例如水杨酸途径)的激活,从而增强了对氧化应激的耐受性。

MED8是EAL4表达的负向调控子的鉴定
A. 用10 mM H2O2处理6小时前后的ProEAL4:LUC和ceal5(MED8突变)幼苗在多孔板中生长的10天龄幼苗的照片(左图),PSII效率Fv'/ Fm'(中图)和发光图像(右图) 持续6小时。

med8的突变激活H2O2依赖的细胞死亡反应


通过在SA或JA生物合成和信号传导中引入二级突变,对med8氧化应激耐性表型进行遗传调控。
A. 在具有或不具有25 nM MV的½MS培养基上生长的3周龄Col-0,med8和med8双突变植物的表型。SA或JA生物合成和信号传导的突变是通过在med8背景中引入sid2和npr1或aos,jar1和myc2突变来实现的。
E. 3周龄的土壤生长的Col-0,cat2,cat2 med8和cat2 med8 sid2植物的表型。长时间光照:16 h光照/ 8 h黑暗);光照强度:200μmol.m-2.s-1。

NOT2蛋白与MED8相关联,并调节氧化应激表型和EAL4表达。
C. 在对照组(1 / 2MS培养基)和MV胁迫组(25 nM MV)条件下萌芽和生长的3周龄植物。明场(上面)和颜色编码的Fv'/ Fm'(下面)图像。
*:本文献中使用了德国伯托的植物活体影像系统Berthold NightShade。
文章链接:https://academic.oup.com/plcell/advance-article/doi/10.1093/plcell/koab079/6169542?login=true
